Rhodophyta also have notoriously few genes for multicellular eukaryotes, possibly due to a genetic bottleneck they went through somewhere in their evolution.
It is unclear whether this is because their ancestors were extremophiles or because of competition with cyanobacteria and Chlorophytes or a combination of both.
But this lack of genes is has confined them to a mostly marine, aquatic environment, with only 5% of red algae living in fresh water and no terrestrial species except for two single celled cave dwellers.
On the distant world of Eryobis however, Rhodophytes are the only multicellular autotrophic organisms introduced. Here they enjoyed competition free seas and lakes and rapidly diversified in the beginning.
While adaptive speciation played a major role, the engine of their evolution came from above.
In the beginning, Eryobis only had a very weak ozone layer and while it had an atmosphere that had broken the moon free of tidal locking, it was not capable of sustaining terrestrial life from Earth, which it still doesn't.
Besides cosmic radiation and lots of solar radiation reaching the ground, the moon was also bombarded with synchotron radiation emitted form Antæios, the planet that Eryobis orbits.
Eryobis' own magnetosphere was enough to keep the radiation from killing everything instantly, although the ionizing properties of the radiation caused a very high mutation rate, aiding the speciation.
For the Rhodophytes, this was not a bad thing, as the radiation induces several polyploidization events in multiple lineages. In such an event, the entire genome of a species is duplicated, which can lead to greater adaptability, flexibility and diversity.
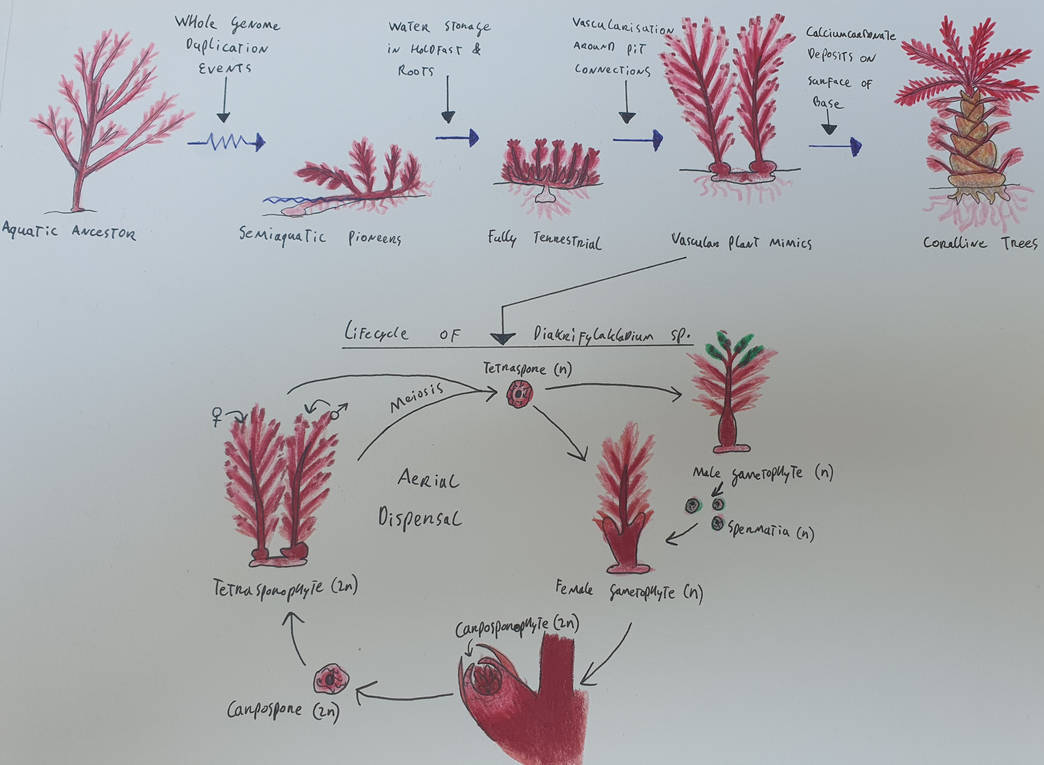
Where the red algae lagged behind on Earth, they made up for on Eryobis. With genes to spare, they evolved ways that allowed them to colonise many aquatic habitats and even conquer land.
The first adaptation for moving out of the water was the ability to retain moisture in the blades (leaves but actually not) when exposed to air, while the holdfast remained in the water close by. Small projections from the main stem that grew downwards evolved into roots.
As seasons are very extreme on Eryobis, this semiaquatic lifestyle was not going to last long. So they evolved a way to keep vital nutrients and water inside de holdfast, which would be buried in the soil, so that it could act similar to the hump of a camel, or in lesser extent as a tuber. After this adaptation, the terrestrial red plants looked more or less like mosses for quite some time before the next big thing evolved.
Almost all member of the Rhodophyta have incomplete cytokinesis were a small pore is left between the mother and daughter cell. These are known as pit connections, and they often have stop in them known as a pit plug. It still unclear exactly what purpose these features serve, although they may act as structural reinforcement.
In the terrestrial Rhodophytes, the next big adaptation was the evolution of vein-like structures around de pit connections of the cells. The flow of water and nutrients pressurized the cells, which caused the plants to straighten. These vascular plant mimics are the most successful group of land plants on Eryobis, having spread to almost every corner of the moon. They were much taller than their non vascular ancestors, but were still limited in size by the availability of water, with the biggest not growing much taller than 1.5 meters (5 foot). Under severe predatory pressure however, some evolved a hard exoskeleton made of calciumcarbonate deposits strengthened with cellulose. Within only a few million years the skeletons of these coralline plants allowed them to grow far bigger than any Rhodophyte had ever been and massive forest were formed around equatorial regions.
Illustrated below is the simplified life cycle of the vascular plant mimic genus Diakrifylakladium. Rhodophytes have an unusually complex reproductive strategy that allows for a lot of genetic diversity within a species.
The 'main' phase of Diakriflykladium is the tetrasporophyte. In this phase the plant has diploid (2n) genes and it grows two feather like stalks. One of these stalks will only produce male tetrapsores and the other will only produce female ones which will both be released into the air. The tetraspores are haploid (n) and will grow into either a male or a female gametophyte (n) depending on the genetic material. The male gametophyte will produce spermatia (n) in their spermatangia. The spermatia are released into the air where they will hopefully find a female gametophyte, if that happens fertilization occurs in the female's carpogonium. In the carpogonium develops the diploid carposporophyte (2n) which will grow its own spore growing organs called carposporangia. The carposporangia release diploid carpospores (2n) into the air and when they settle, they will grow into the bisexual tetrasporophyte and the whole cycle begins again.



No comments:
Post a Comment